肠道菌群在结直肠癌中的作用及其机制
杨永涛,屈昌民,李连勇*,中国人民解放军第306 医院消化内科
诸多研究表明肠道细菌是结直肠癌( CRC) 的致病因素。目前研究已发现可增加CRC 风险的相关菌属。肠道细菌可通过对肠黏膜上皮细胞作用,如影响DNA 完整性、调节免疫反应、诱导炎症反应、促进细胞增殖、改变干细胞动力学等,促进CRC 发生。调整肠道菌群有望成为预防或治疗CRC 的手段。
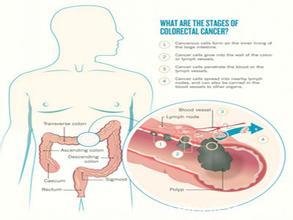
微生物参与多种肿瘤的发生、发展,至少15% 的肿瘤由感染因素引起。结直肠癌( colorectal cancer,CRC) 是世界第三大恶性肿瘤,息肉-腺瘤-癌是CRC 的主要演变模式。大部分CRC 呈散发性,提示环境因素而非遗传因素是CRC 的主要诱因。结肠中的细菌含量约为小肠中的1010 倍,同时结肠癌的发生率约为小肠癌的12 倍,说明肠道菌群在肠道肿瘤中发挥重要作用。既往研究表明西方化饮食方式与CRC 的发生密切相关,饮食结构引起的肠道菌群变化可能介导了CRC 发生。肠道菌群可通过产生代谢产物,诱导黏膜免疫反应,调节细胞增殖等方式促进CRC 发生。
一、肠道菌群的组成和功能
肠道菌群是由超过1 000 余种细菌组成的复杂微生物群落。由于不同节段肠管的传输时间、pH 值、氧含量、营养成分、分泌功能等的不同,细菌在肠道不同节段的定植存在差异。肠道中的细菌群落主要集中于结直肠( 每克肠内容物约含1012 个细菌) ,主要为厌氧的硬壁菌门和拟杆菌门,此外亦包括放线菌门、变形菌门、疣微菌门等。尽管超过80%的肠道细菌不能体外培养,但DNA 序列分析使肠道菌群的分类和定量成为可能。通过提取粪便细菌DNA 进行全基因组测序发现,虽然不同个体间肠道菌群存在差别,但成年后个体肠道菌群可保持相对稳定。对欧洲健康志愿者粪便细菌DNA 测序发现,根据优势菌属的种类,可将肠道菌群分为三个“肠型”,即拟杆菌属型、普氏菌属型和瘤胃球菌属型,对日本人群的研究亦得到相似结果。肠道菌群的组成可受饮食因素影响,饮食结构改变可使肠道菌群在数天内发生改变[4]。碳水化合物的摄入可导致厚壁菌门和放线菌门细菌组成发生重大变化。低碳水化合物的食物能够使厚壁菌门中的真杆菌属和放线菌门中的双歧杆菌属等产丁酸盐细菌减少。而以植物为主的饮食( 高纤维素、低脂肪、低蛋白质) 相对于以动物为主的饮食( 低纤维素、高脂肪、高蛋白质) 会产生更广泛的肠道菌群变化。动物性饮食会使拟杆菌门细菌增加,放线菌门细菌减少[8]。高蛋白质和高脂肪饮食可促进包括普通脱硫弧菌在内的硫酸盐还原菌生长,并产生过量硫化氢。高脂肪饮食可产生胆汁酸,后者是表达7α 脱羟基酶细菌的适合底物,这些细菌可将初级胆汁酸转变为具有潜在致癌作用的次级胆汁酸。正常肠道菌群与人体间存在共生关系,细菌在促进人体免疫系统发育、成熟,参与形成黏膜屏障,防止病原菌定植,解毒环境中致癌因子等方面发挥重要作用。另一方面,人体的遗传背景、免疫系统以及激素水平等因素可影响肠道菌群组成,因此由人体遗传背景和生活方式差异造成的肠道菌群差异可能是导致不同个体对肥胖、哮喘、自身免疫性疾病、CRC 等细菌相关性疾病易感性不同的原因。
二、肠道菌群在CRC 发生中的作用
高纤维素尤其是谷物和粗粮的摄入可降低CRC 风险。食物中的膳食纤维仅小部分在小肠吸收,约95% 未被消化的纤维到达结肠,在肠道菌群作用下被分解或转换,提示肠道菌群在CRC 发生中发挥重要作用。诸多证据表明肠道菌群是CRC 的重要致病因素。
1. 动物研究: APCmin/ + 小鼠是广泛应用的CRC 动物模型,其携带APC 基因杂合突变,能自发形成肠道肿瘤。Son等的研究显示,与正常环境下饲养的APCmin/ + 小鼠相比,无菌条件下饲养的APCmin/ + 小鼠肠道腺瘤的数目和体积明显减少。同样,无菌环境下饲养,经氧化偶氮甲烷( AOM) 和葡聚糖硫酸钠( DSS) 诱导的白细胞介素( IL) -10 - / -、Tcrb /p53 - / -、Gpx1 /Gpx2 - / - 和Tgfb1 /Rag2 - / - 小鼠模型较有菌环境下饲养的具有相同遗传背景的小鼠模型产生肠道腺瘤的数量明显减少,证实肠道微生物参与了CRC 的发生。Nagamine 等的研究发现,肝螺杆菌可促进AOM 诱导IL-10 - / - 小鼠发生结肠炎相关肿瘤,而拟杆菌可抑制IL-10 - / -小鼠肠道肿瘤形成,说明相同遗传背景下不同细菌在结肠癌发生中的作用不同。与无菌动物相比,移植人体肠道菌群小鼠的肠黏膜上皮细胞增殖更活跃,抗菌药物可降低小鼠结肠炎相关CRC 的发生风险,而通过共同饲养或菌群移植的方式可将结肠炎相关结肠癌传递给遗传易患子代小鼠。上述研究说明肠道菌群在CRC 的发生中发挥重要作用。
2. 临床研究: 1977 年Savage DC 等报道了与CRC 相关的13 种细菌,但由于样本量较小以及CRC 诊断技术的限制,其结果并未获得广泛认可。随后有研究发现幽门螺杆菌、黏附侵袭性大肠杆菌与CRC 相关。肠道菌群的组成、分布以及代谢改变可使结肠组织向炎症、异型增生以及肿瘤发展。Marchesi 等发现,CRC 患者粪便中用于构建系统发育树的核心基因发生显著变化。McCoy 等的研究表明,与健康人群相比,腺瘤患者肠黏膜细菌多样性增加,粪便拟杆菌属数量增加,产丁酸盐细菌数量减少,具有潜在致病性的具核梭杆菌属数量增加。然而,Pagnini 等的研究发现,黏附于结肠腺瘤组织上的细菌数量较正常组织少20 倍,可能是由于腺瘤组织相对正常组织分泌更多的α-防御素所致,但并无更多证据支持此种假设,有待进一步研究证实。

三、菌群失调在CRC 发生中的作用机制
1.对于肠道菌群引起CRC 的机制,有学者提出起始菌-过路菌模型,该模型指出致病菌通过引起肠上皮细胞DNA损伤诱发CRC,CRC 亦可进一步引起肠道环境改变使机会致病菌增殖、起始菌减少。1. 肠道菌群对肠上皮细胞致畸作用: 正常肠黏膜上皮细胞沿隐窝-绒毛轴进行快速更新,由于位于肠黏膜隐窝底的干细胞及其前体细胞突变易累积额外突变,因此CRC 被认为是由基因突变所致。研究显示部分肠道致病菌可引起肠上皮细胞基因突变。脆弱类杆菌可诱导结肠上皮细胞表达精胺氧化酶,产生反应性氧自由基,引起DNA 损伤。粪肠球菌可产生细胞外超氧化物和过氧化氢,损伤结肠上皮细胞DNA[19]。具核梭杆菌可产生黏附分子FadA,后者不仅可帮助细菌黏附至肿瘤细胞,亦可结合E-钙黏蛋白,活化β-连环蛋白( β-catenin) 信号通路,从而上调转录因子、癌基因以及炎症相关基因,最终诱发CRC 细胞增殖,促进肿瘤的生长。部分肠致病性大肠杆菌可编码聚酮肽基因毒素“Colibactin”,使宿主细胞DNA 双链断裂,激活DNA 损伤信号级联反应,导致慢性有丝分裂和染色体畸变以及基因突变频率增加,从而引发肿瘤。基因敲除聚酮化合物合成的致病岛后,大肠杆菌引起的炎症水平无显著改变,但肿瘤细胞侵袭能力有所减弱,提示细菌合成的聚酮化合物可促进肿瘤发生。在CRC 发病早期阶段,β-catenin 和APC 基因突变可导致结肠上皮屏障功能缺失,导致微生物产物转运至肿瘤微环境,此过程可使产生IL-23 的髓系细胞活化,产生大量细胞因子促进肿瘤生长。
2. 细菌代谢产物对黏膜上皮细胞的作用: 纤维饮食中的碳水化合物经细菌分解发酵后,可产生包括丁酸、丙酸、乙酸在内的短链脂肪酸( SCFAs) ,其中丁酸盐具有抗肿瘤特性,可降低CRC 发病率。梭菌Ⅳ群中的柔嫩梭菌属和梭菌ⅩⅣa 群中的真杆菌属是人体结肠内产生丁酸盐的主要细菌。SCFAs 可与结肠上皮细胞和免疫细胞表面的G 蛋白耦联受体( GPCRs) 结合并被转运至细胞内,细胞内的SCFAs 可抑制组蛋白去乙酰化酶( HDAC) 的活性,导致组蛋白过乙酰化。HDAC 的抑制和GPCRs 介导的信号转导能使结肠调节性T 细胞数量增加,并且诱导抗炎性细胞因子IL-10 等产生。GPCRs 信号转导亦可通过非依赖HDAC 途径抑制核转录因子( NF) -κB 活化,从而引起细胞凋亡。同时,丁酸盐和丙酸盐可通过活化AP-1 信号通路抑制细胞增殖以及选择性诱导细胞凋亡。肠道微生物可参与植物化合物和外源性有毒物质的生物转化,这些物质在肠道吸收后,在肝脏与葡萄糖醛酸等物质结合,通过胆汁排入肠道。肠道细菌中的β 葡萄糖醛酸酶可将葡萄糖醛酸转换为苷元重新吸收,延长了上述物质在人体内的存留时间。已有研究[26]证实,高β 葡萄糖醛酸酶活性可增加CRC 风险,提示肠道细菌可通过β 葡萄糖醛酸酶参与CRC 发生。高蛋白饮食会使细菌水解发酵肠道内过量的蛋白,产生具有致炎和致癌作用的酚、氨、支链短链脂肪酸和其他含氮代谢产物。高蛋白和高脂肪饮食可促进硫酸盐还原菌生长,产生大量具有基因毒性的硫化氢。未经小肠重吸收的部分初级胆汁酸在结肠内经细菌作用后脱去甘氨酸和牛磺酸残基,再经过7α 脱羟基细菌作用,生成包括脱氧胆酸和石胆酸在内的次级胆汁酸,后者通过产生具有基因毒性的活性氧和活性氮,参与CRC 的发生。此外,过量摄入乙醇亦是诱发CRC 的危险因素,其氧化产物乙醛具有较强的毒性和致癌作用,可导致细胞从DNA 损伤至叶酸降解等不同程度的损害。研究表明,口腔微生物有助于乙醇转化为乙醛,提示肠道微生物亦可能通过此途径参与CRC 的发生。
3. 细菌诱导的炎症反应参与肿瘤发生: 微生物的脂多糖、鞭毛蛋白、肽聚糖以及其他微生物相关分子模式( MAMPs) 可被天然免疫系统,如NOD 样受体( NLRs) 、Toll样受体( TLRs) 和视黄酸诱导基因-Ⅰ样受体在内的模式分子受体( PRRs) 识别。天然免疫系统的激活可调节细胞增殖、黏膜屏障功能以及多种细胞炎症通路[10]。研究表明,与腺瘤和正常组织相比,癌组织上皮细胞TLRs 表达增加。另一项关于癌组织cDNA 微阵列研究的meta 分析发现,癌组织中NLRs 信号通路相关基因表达明显异常。PRRs通路相关基因修饰的模型小鼠可用于研究肠道细菌致CRC的机制,研究发现先天免疫途径介导了细菌或化合物引
起的肠上皮细胞慢性炎症反应。慢性炎症是诱导CRC 发生的一个主要危险因素,炎症可募集和释放肿瘤坏死因子( TNF) -α、IL-6、IL-1 等细胞因子,后者通过活化NF-κB 等途径参与CRC 的发生。TLR4 基因缺失小鼠发生CRC的风险低,而过表达TLR4 可增加AOM 和DSS 诱导结肠炎相关肿瘤的发生风险[28]。MyD88 是传导TLRs 信号的接头蛋白,MyD88 - / - 小鼠经AOM 诱导发生CRC 的风险较野生型小鼠降低,但AOM 与DSS 联合诱导发生结肠炎相关CRC的风险增加,此可能与髓系细胞IL-18 产生缺失有关。研究显示,定向敲除小鼠肠道上皮细胞APC 等位基因后,细菌可诱发小鼠发生CRC,且肿瘤组织中IL-23 和IL-17 表达增加,而给予抗菌药物治疗或敲除MyD88 基因后, IL-23和IL-17 水平以及肿瘤生长速度均下降。Salcedo 等的研究亦指出,MyD88 - / - APCmin/ + 小鼠较APCmin/ + 小鼠产生结肠肿瘤的数目减少,说明在APC 基因突变的条件下,细菌介导的信号可促进肿瘤发生。此外,有研究发现MyD88 可通过活化ERK 信号通路,抑制β-catenin 降解,从而促进肿瘤发生。然而,亦有研究[28]指出TLRs - / - 小鼠较野生型小鼠易产生胃肠道肿瘤,相关机制仍需深入研究。
四、展望
虽然CRC 具有遗传倾向,但环境因素在CRC 发生、发展中占据主导地位,诸多研究表明肠道细菌是CRC 的致病因素。随着科学技术的发展,肠道菌群的组成、功能、稳定性、宿主特异性、功能基因以及宏基因组成为研究热点。目前研究已发现可增加CRC 风险的相关菌属。肠道细菌可通过对肠黏膜上皮细胞作用,如影响DNA 完整性、调节免疫反应、诱导炎症反应、促进细胞增殖、改变干细胞动力学等,促进CRC 发生。调整肠道菌群有望成为预防或治疗CRC 的手段。此外,肠道菌群对外源性化合物的代谢会影响CRC治疗疗效,了解肠道菌群组成和代谢有助于治疗方案和监控策略的选择,从而提高治疗疗效,此亦为使用表达特定基因或生产特定代谢产物的细菌治疗或预防CRC 提供可能。