第122期:重置“夜猫子”的晚睡时间对心理健康和表现有积极的影响
本期由姝儿分享 Elise R等人2019年发表在 Sleep Medicine上的题为“Resetting the late timing of ‘night owls’ has a positive impact on mental health and performance的文章。
本文选择了一组“夜猫子”(即睡眠和清醒模式极其晚的人),并试图在现实环境中使用简单、实用的非药物干预措施,提前改变他们习惯性的晚睡时间。我们表明,通过使用这种干预,我们可以:
提前两个小时左右入睡。
降低自我报告的抑郁和压力等级。
减少早晨的困倦。
显著改善认知和身体表现的简单指标。
01摘要
按照我们内在的生物节律生活和我们的外部环境之间存在冲突,干扰会对健康和表现产生负面影响。这通常在轮班工作和时差反应中有记录,但“社会规范”,如典型的工作时间,会给“夜猫子”带来深远的问题,这些人的内部生物时间使他们倾向于遵循异常晚的睡眠-觉醒周期。夜猫子也与健康问题、情绪障碍、表现不佳和死亡率增加有关。这项研究使用了一项随机对照试验设计,旨在将夜猫子的晚睡时间提前到更早的时间(相位提前),在现实环境中使用非药物的、实用的干预措施。这些干预措施针对光照(通过更早的醒来/睡眠时间)、固定的用餐时间、咖啡因摄入和锻炼。总体而言,参与者通过活动描记法和昼夜节律相位标记(昏暗的褪黑激素发作和皮质醇觉醒反应的峰值时间)测量,显示睡眠/觉醒时间显著提前约2小时,同时对睡眠持续时间没有不利影响。
重要的是,相位提前伴随着自我报告的抑郁和压力的显著改善,以及在典型的“次优”早晨时间内认知(反应时间)和身体(握力)表现指标的改善。本文的发现提出了一种新的策略,将时钟计时转向一种更符合社会需求的模式,这种模式可以显著改善现实世界中的表现、心理健康和睡眠时间等要素。
02介绍
通常很少考虑睡眠和昼夜节律紊乱对社会对我们典型工作日组织的态度的影响。睡眠/唤醒系统的干扰损害了日常功能,导致健康下降,这种情况很普遍,大约三分之二的英国成年人(67%)报告了某种睡眠问题。
众所周知,睡眠受限和昼夜节律紊乱会导致许多生理过程的变化,如内分泌调节和核心体温(CBT),并与各种健康问题有关,包括情绪障碍、发病率和死亡率增加以及认知和身体表现下降。昼夜节律和睡眠/觉醒过程的中断给社会带来了巨大的经济负担,主要表现为生产力损失、缺勤和表现不佳,并增加了职业事故的风险。影响这些结果的一个主要因素是缺乏对易受睡眠中断和昼夜节律失调影响的个体差异的认识,以及缺乏对个体的昼夜节律时间可能与正常工作日09:00-17:00不一致的程度的认识。
生物节律的个体差异受生理、遗传和行为因素的影响。这些差异允许根据个体的昼夜节律时间进行分类,特别是早期和晚期时间通常被称为“云雀”和“夜猫子”(在本研究中被称为早期和晚期昼夜节律表型,ECP/LCP)。在最极端的情况下,这些差异可能导致昼夜节律睡眠-觉醒障碍(CRSWDs)、高级睡眠-觉醒期障碍(ASWPD)和延迟睡眠-觉醒期障碍(DSWPD)的临床诊断,这三种疾病分别在老年和年轻受试者中更为普遍。在健康人群中,这些临床疾病在极端昼夜节律表型机制方面的重叠程度仍然未知。
DSWPD通常与抑郁等情绪障碍有关,这一群体也往往受到社会因素的限制,如缩短睡眠的工作/学校惯例,导致“睡眠债”的积累。这会导致白天过度嗜睡和认知功能受损。虽然需要临床评估来诊断糖尿病周围神经病变,它的许多症状都与“夜猫子”有关。基于睡眠/唤醒时间的延迟、昏暗光线下褪黑激素发作的延迟(DLMO)和/或睡眠稳态的缺陷对睡眠障碍进行分类。LCPs与抑郁得分较高、早晨认知能力下降、白天过度嗜睡以及发病率和死亡率风险增加有关。认知和身体表现指标的昼夜变化也显示出昼夜表型之间的 差异,而LCP通常难以适应传统的工作时间。由于给定人群中约50%属于“晚型”(8:18后醒来),人们可能会认为这些人因昼夜节律计时延迟而受到损害,并可能因转向更早的模式而受益。
重置生物钟可以通过行为方法、药理学方法或两者的结合来实现。人类的昼夜节律系统对光最敏感,这使得睡眠/觉醒活动和生理适应24小时的光暗周期。因此,光或缺乏光是试图通过一种叫做光夹带的过程来重置生物钟的主要目标。已有研究表明,强光会根据光照时间和持续时间改变昼夜节律相位(相位响应曲线)。清晨阶段的暴露会使昼夜节律系统提前,导致DLMO高峰提前,睡眠开始提前。相反,在生物之夜暴露在光线下会产生相位延迟,就像后来的DLMO所显示的那样。
还研究了非光形式的夹带,试图改变昼夜节律相位。这些行为目标,即非药物干预,包括改变睡眠/觉醒周期、定时体育锻炼和定时进食。定时喂食已被证明可以在不影响SCN时钟的情况下改变小鼠的外周时钟。此外,定时进食被证明可以调节外周代谢节律,进餐时间延迟5小时会延迟血浆葡萄糖和脂肪PER2时钟基因表达的节律。另一种已经被探索过的昼夜节律疗法是有针对性的体育锻炼。定时运动可以改变核心体温和褪黑素的节律。最近的一篇论文进一步支持了这些发现,表明上午和下午早些时候的锻炼会导致相位提前,而晚上的锻炼会导致相位延迟。我们社会的大多数人都有严格的工作和上学时间,要求在09:00到17:00之间出勤。尽管有这些传统的强加的社会时钟要求,但通过允许工作时间的灵活性,以及试图改变学校的开学时间以适应青少年众所周知的迟到生物钟,已经朝着理解生物约束的方向发生了一些转变。然而,尽管意识到了后果,但要直接转化研究成果并影响我们快速发展的“全天候”社会的变化,还有很长的路要走。虽然使用这些方法中的一些来尝试相位提前(更早地改变时钟)以前已经在实验室研究中显示过,但是缺乏实地研究。此外,在现实环境中,还没有尝试调查对心理健康的影响和表现的昼夜变化。在这里,我们提出了一种新的“夜猫子”干预策略,其中许多人患有慢性昼夜节律失调或睡眠稳态中断。通过简单、实用的生活方式改变,我们旨在逐步提前睡眠/醒来时间、DLMO和皮质醇觉醒反应峰值时间。我们假设,如果提前一个阶段,这将改善心理健康(抑郁、焦虑和压力)的自测指标,并提前改变最佳表现的时间,从而改善一天中非最佳时间的认知(反应时间)和身体(握力)表现的简单指标。
03方法
受试者
该研究获得了伯明翰大学研究伦理委员会的良好伦理意见,根据赫尔辛基宣言进行,参与者在参与前给予书面知情同意。共有178人完成了慕尼黑计时问卷(MCTQ,纸质版),以计算自由日的修正睡眠时间。使用年龄和性别匹配的MCTQ数据库分类为晚期时间类型的参与者被邀请参加研究(n = 49)。通过自我报告,对个人进行了睡眠或神经障碍诊断筛查,并且没有服用任何影响睡眠、褪黑激素和皮质醇节律的药物。共有29人同意参与研究,其中5人因病史被排除在外,2人在研究开始前退出。最终样本由22名健康个体组成(15名女性,年龄21.3±3.3岁,MSF sc 06:52 00:17 h)。该研究采用随机对照试验设计,在2016年4月至6月期间对每位参与者进行了为期六周的研究(日出时间范围为06:42小时至04:40小时,日落时间范围为19:41小时至21:32小时,北纬52° 29 ' 22.0956 "。在研究开始时,参与者被随机分配到实验组(n = 12,9名女性)或对照组(n = 10,6名女性)。两周的适应被用来评估习惯性睡眠模式,使用活动描记法和收集基线(干预前)的问卷数据。在此期间之后,参与者被要求在14:00、20:00和08:00前往实验室进行测试之前,提供其家庭环境中褪黑激素和皮质醇的唾液样本(详情如下)。为了模拟“真实世界”的环境,参与者可以在测试期间离开实验室。然后给参与者一个接下来三周的时间表(干预),然后返回重复所有的测试、生理取样和问卷(图1)。参与者在干预前和干预后的同一天完成测试。在补充表1中可以找到实验组和对照组的参与者数据预干预的概要细节,以确认准确的匹配。
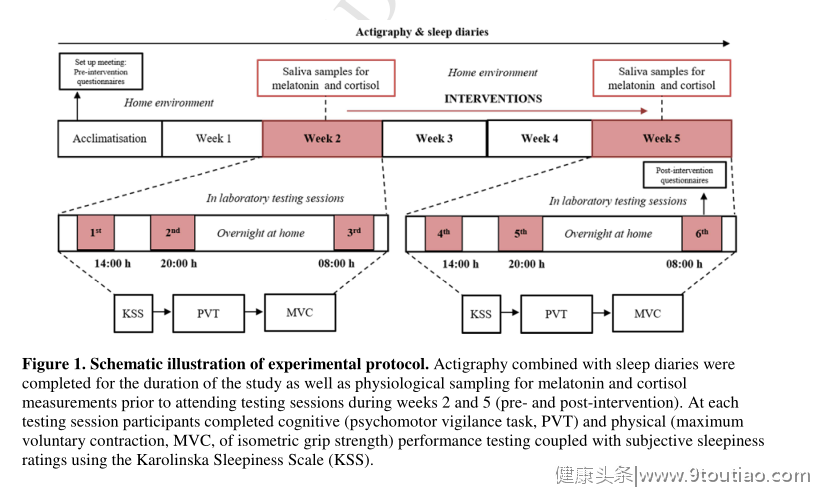
非药物干预
在最后的干预前测试阶段,实验组被给予为期三周的干预时间表。这些干预措施遵循标准的睡眠卫生建议,并针对适当的定时光照、睡眠、膳食、咖啡因和锻炼(总结在表1中)。对照组只给了安慰剂一条“每天在同一时间吃午餐”的指令,假设睡眠时间没有差异,因此对昼夜节律期没有影响。通过自我报告监测干预的遵守情况。干预前后的进餐时间作为饮食问卷的一部分进行收集。午睡的时间通过每天的睡眠日记来监控。在研究结束时进行了一次反馈问卷调查,参与者被问及他们是否遵守了干预时间表,范围从0(完全没有)到10(完全遵守)。在每一次测试中,参与者都要回答一份在线问卷,以记录测试前后的外部变量,如咖啡因摄入量、锻炼和用餐时间。

所有参与者都接受了如何在家庭环境中按照严格的协议收集唾液样本的培训。在取样期间,参与者被要求避免清洁牙齿、饮用含咖啡因的饮料、酒精饮料或任何含有人工色素的饮料。向每个人提供一份样本记录收集表,以便报告提供样本的准确时间,并报告可能影响采样周期的任何因素,例如暴露在光线下、采样中断。参与者在干预前(第2周)和干预后(第5周)的一个上午和一个晚上提供唾液样本。从习惯性就寝前3至4小时到习惯性就寝后1小时,每30分钟收集一次褪黑激素样本,同时坐在昏暗的照明条件下,即没有顶灯、没有电子设备和窗帘关闭。皮质醇的早晨样本是在从醒来开始的3小时内收集的(前五个样本每15分钟收集一次,其余四个样本每30分钟收集一次)。记录每个个体的准确采样时间。使用碘125放射性标记的示踪剂和固相分离进行人唾液中褪黑激素和皮质醇的放射免疫测定。
使用个体干预前浓度值的平均值加上平均值的两个标准偏差,用线性响应函数计算个体DLMOs。取样期间记录的最高皮质醇浓度时间被用作皮质醇觉醒反应峰值的指标。由于样本不足或受污染,无法计算实验组的三名受试者和对照组的五名受试者的配对DLMO值(干预前和干预后)。
行为数据
睡眠分析:在整个研究期间(第1-5周),将活动描记器(活动观察灯,2006,剑桥神经技术有限公司)与日常睡眠日记相结合,戴在非主导的手腕上,以监测家庭环境中的活动睡眠和休息活动模式(1分钟时期),并用制造商的软件进行分析。由于装置佩戴不当,来自两个人(一个在实验组,一个在对照组)的活动数据不可用。调查问卷:每个参与者在干预前的设置会议中完成一套调查问卷,并在最后的测试会议(干预后)结束时重复。调查问卷包括心理健康问卷、纸质版、爱普沃斯嗜睡量表(ess)、匹兹堡睡眠质量指数(PSQI)、情绪状态概况(POMS)、抑郁、焦虑和压力量表(DASS)和饮食问卷。由于问卷填写不充分,POMS有三人的结果没有记录,两人的饮食调查问卷和两人的饮食调查问卷没有记录。嗜睡:白天主观嗜睡,使用卡罗林斯卡嗜睡量表(KSS)测量,在认知和身体任务执行前的每个测试阶段进行评估。反应时间:认知测试包括两分钟的视觉精神运动警戒任务(PVT)。PVT是在一台带有标准键盘和鼠标的台式计算机上进行的。在整个研究过程中,每个参与者和每个测试环节都使用了相同的设置。参与者还在适应阶段进行了三次试验测试,以熟悉设置并最大限度地减少学习效果。记录每个试验的毫秒数,然后取试验次数的平均响应时间。
握力:为了获得物理性能的简单测量,使用电子手动测力计(EH101,凯美瑞)对等长握力进行6秒钟最大自主收缩测试。参与者站立时肘部伸展至180度,并以旋前的姿势用他们的优势手施加尽可能多的握力。原始分数以千克为单位记录。完成了三项试验,每项试验之间休息两分钟,在随后的分析中使用最高记录值。由于动机对表现的影响,一套脚本被用来激励参与者。
统计分析
在GraphPad Prism(7.00版)中进行统计比较,使用线性回归分析和双向重复测量方差分析,事后检验针对多重比较进行校正,添加干预组(实验组/对照组)、评估期(干预前与干预后)或一天中的时间(08:00小时、14:00小时和20:00小时)作为因素。使用二次多项式回归曲线绘制了表现和嗜睡变量的日变化。由于数据收集发生在14:00到08:00,模型受限于此时间段。
性能测量的原始分数(PVT的反应时间以毫秒为单位,MVC测试的握力以千克为单位)通过转换为相对于每个人最佳表现时间的百分比进行标准化。例如,记录了最快反应时间和最强握力的测试时段被指定为该参与者的100%。随后的分数是相对于此计算的。更高的百分比总是与获得更好的性能(更快的反应时间和更强的抓地力)有关。这是为了能够以标准化的方式量化个体之间以及不同绩效指标之间的日变化(Facer-Childs等人,2018年)。这些数据分别在干预前和干预后相对于每个个体进行标准化。测试统计给出了一个重要的数字。显著性水平显示为ns =不显著,p < 0.05 = *,p < 0.01 = **,p < 0.001 = ***和p < 0.0001 = ***。除非另有说明,否则数值表示为平均值的平均标准误差(年龄和体重指数值以标准偏差给出)。除了显著性被确定为小于0.0001(在这种情况下,报告p < 0.0001)之外,精确的p值被赋予两个显著数字。08:00 h测试描述为上午,14:00 h为下午,20:00 h为晚上。反应时间(使用PVT测量)将被称为注意力认知表现的简单指数,等长握力(使用MVC测试测量)将被称为 身体表现的简单指数。
04结果
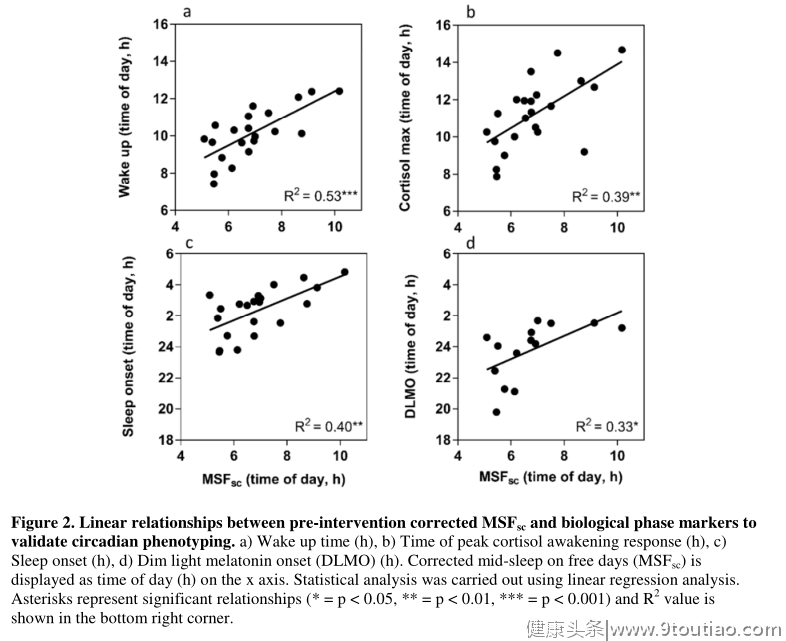
为了确认根据下面讨论的变量范围,研究组是均匀匹配的,所有数据在干预前进行了初步比较,在测量的任何参数中没有发现显著差异。实验组和对照组年龄相近(21.7±2.8和20.9±3.9岁),BMI(22.9±3.2和22.6±2.1)和MSF sc (07:15 h 291 00:27和06:02 h 00:14)。在基线时(干预前),观察到睡眠质量分数与唤醒时间(R 2 = 0.53,F = 21.21,p = 0.0002)、睡眠开始时间(R 2 = 0.40,F = 12.69,p = 0.0021)、皮质醇觉醒反应的峰值时间(R 2 = 0.39,F = 12.65,p = 0.002)和DLMO (R 2 = 0.33,F = 5.98,p = 0.001)之间存在显著的线性关系这些结果支持并验证了参与者通过活动分析和生物相标记被分类为LCPs的观点,这些活动分析和生物相标记是在最初从MCTQ鉴定为晚期年代型之后进行的。
05坚持干预
总体而言,实验组在反馈问卷中报告的干预措施的遵守率平均为7.8±0.7。坚持针对睡眠/觉醒和饮食变量的干预措施(通过活动描记法和饮食问卷监测)随着时间的推移而得到证实。使用自我报告的睡眠日记证实了16:00后避免小睡。在晚间测试环节,来自在线问卷的332个结果证实了实验组干预后自我报告的咖啡因摄入、锻炼和最后一餐的时间提前。平均自我报告的咖啡因摄入量在干预后平均提前4小时(干预前5.9±1.7小时,干预后10.3±1.5小时),这意味着这一时间从下午2点提前到上午10点。自锻炼以来的平均自我报告小时数从干预前的6.8±1.7小时增加到干预后的7.8±1.8小时,自最后一餐以来的小时数从干预前的2.4±0.4小时增加到干预后的3.8±0.8小时。相比之下,相对于干预前,对照组的运动时间和用餐时间略有延迟(分别为6.0±1.9小时至4.8±1.8小时和3.5±1.0小时至2.8±0.8小时),咖啡因的使用时间略有提前(从6.1±2.3小时至8.7±2.6小时)。
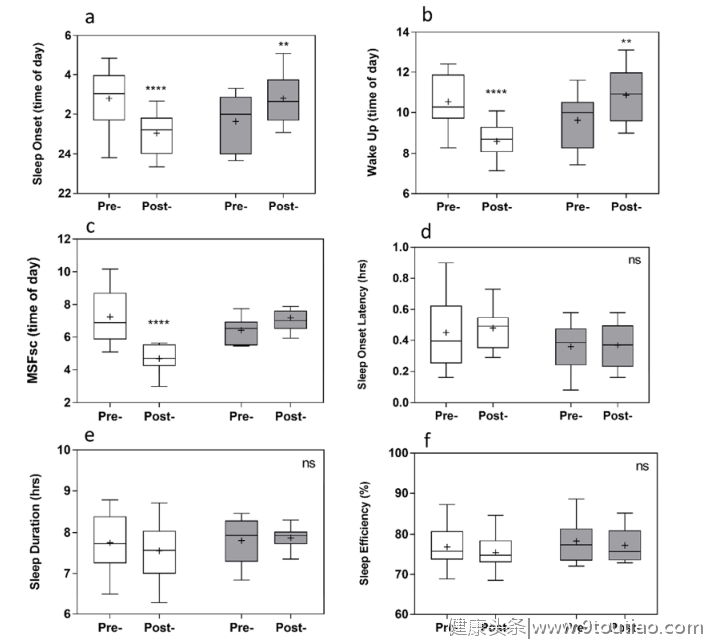
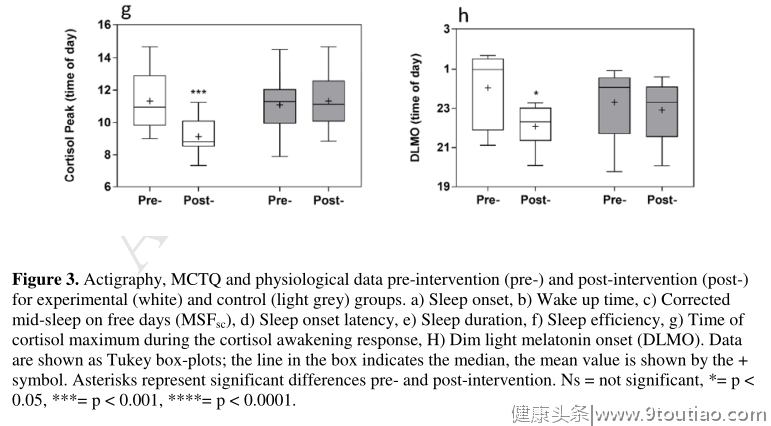
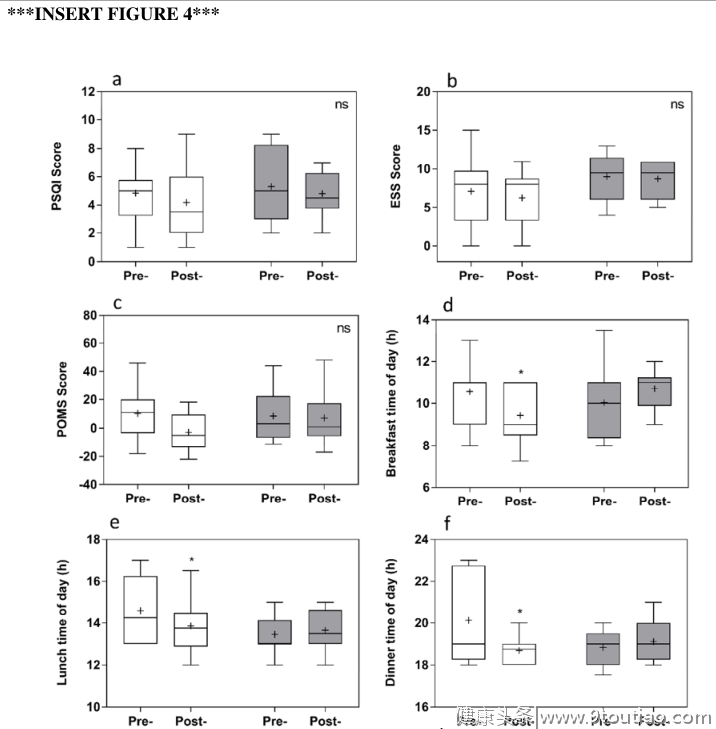
与干预前相比,干预后在实验组中观察到约2小时的明显相位提前,这是通过MCTQ、活动图和昼夜节律相位标记来测量的(图3和表2)。MSF sc显著提前2.57±0.32小时(p < 0.0001)。这一进展得到了行动分析的证实,显示睡眠开始时间显著提前1.73±0.28小时,醒来时间显著提前1.92±0.26小时(均p < 0.0001),睡眠持续时间、睡眠效率或睡眠潜伏期无显著变化。DLMO提前了1.96±0.63小时(p = 0.018),皮质醇觉醒反应峰值时间提前了2.22±0.50小时(p = 0.0005)。相角(睡眠开始和DLMO之间的时间)没有显著变化。与干预前相比,实验组自我报告的平均早餐时间提前了1.11±0.39小时(p = 0.022)。同样,平均自报午餐和晚餐时间也提前了0.75±0.27小时(午餐,p = 0.023)和1.44±0.49h(晚餐,p = 0.021)。与干预前相比,对照组睡眠开始时间显著延迟1.16±0.34小时(p = 0.0067),醒来时间显著延迟1.24±0.32小时(p = 0.0021)。与实验组相比,对照组干预后其他变量无显著差异。
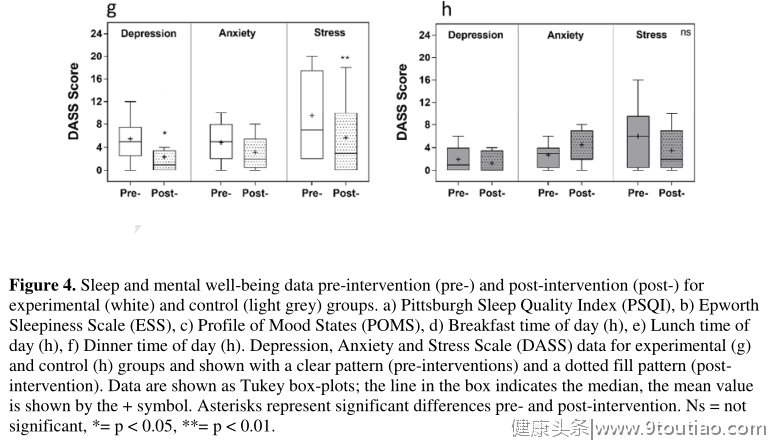
干预对心理健康的影响在实验组进行干预后,抑郁和压力的主观评分显著下降(图4和表2)。总体DASS评分下降8.7±2.4分,从361±19.8降至11.2(干预前)。将DASS分为抑郁、焦虑和压力评分分别显示了干预的显著效果(F(1,11) = 13.28,p = 0.0039),并且显著降低了抑郁和压力因素,但没有降低焦虑(p = 0.37)。抑郁从5.5±1.0降至2.3±1.2(p = 0.025),应激从9.5±2.2降至5.7±1.9(p = 0.0061)。对照组的任何测量参数均无显著差异。在两个研究组中,POMS、PSQI或斯洛文尼亚就业服务局没有观察到显著差异(图4和表2)。
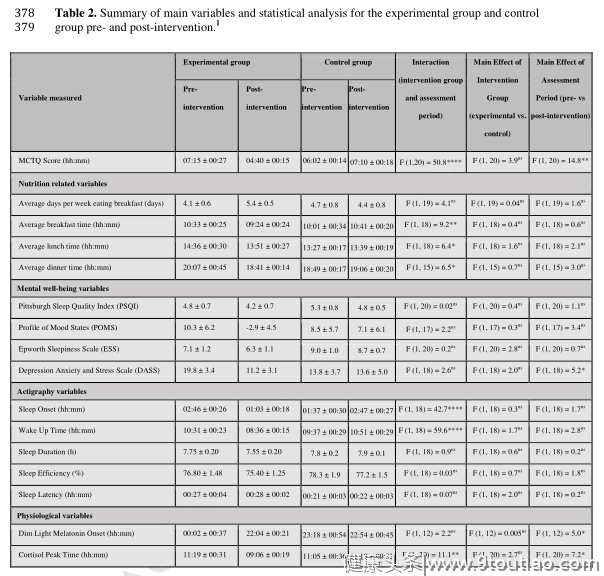
干预对表现和嗜睡的影响使用二阶多项式回归分析,从最佳拟合的日变化曲线中识别出最佳表现和嗜睡时间(图5)。在模型的约束下(08:00 439小时至20:00小时),实验组和对照组在干预前和干预后的08:00时嗜睡程度最高。在干预前测试中,实验组最强的握力出现在20:00测试时段,该测试时段提前到干预后15:21。在对照组中,最大握力的时间从干预后17:12小时延迟到20:00小时。实验组反应时间从20:00小时提前到12:30小时最快,对照组反应时间从15:48小时延迟到19:48小时,PVT也是如此。实验组个体间的表现差异显著降低,而对照组没有显著变化。在干预前测试中,实验组的平均握力变化为14.2%,干预后降至7.2%(p = 0.0024)。反应时间也是如此,平均个体间差异从干预前的13.0%降至干预后的4.4%(p = 0.028)。在实验组(F(2,22) = 3.44,p = 0.049)中,白天时间和干预的显著相互作用被发现,并且白天时间(F(2,22) = 11.41,p = 0.0004)和干预(F(2,11) = 5.36,p = 0.041)的主要影响被发现。干预后,08:00时(4.6±0.6 vs 6.3±0.3)和14:00时(3.6±0.5 vs 4.7±0.5)嗜睡较低,但这些差异仅在08:00时显著(p = 0.0061)。实验组还显示了一天中时间对握力表现的显著主要影响(F(2,22) = 21.73,p < 0.0001),以及干预的显著主要影响(F(1,11) = 4.94,p = 0.048)和交互作用影响(F(2,22) = 9.19,p = 0.0013)。事后测试显示,08:00和14:00时的握力在干预后显著改善(p分别为0.015和0.0075)。对于PVT的表现,有一天的时间的主要影响(F(2,22) = 3.85,p = 0.037),但没有干预。然而,发现相互作用460是显著的(F(2,22) = 7.93,p = 0.0026)。干预后08:00时的反应时间明显快461(p = 0.017),但在14:00时或20:00时没有变化。
在对照组中,发现一天中时间对嗜睡(F(2,18) = 8.86,p = 0.0021)、MVC (F(2,18) = 14.73,p = 0.0002)和PVT (F(2,18) = 3.63,p = 0.048)的表现有显著的主要影响,但对干预或相互作用没有影响。在对照组中,从干预前到干预后,事后测试没有显示任何参数的显著变化。

06讨论
研究人员、临床医生和行业专家正在不断寻找方法,以更好地理解我们如何改善心理健康、福祉和表现。一个似乎经常被忽视的因素是行为的时间安排,例如睡觉、吃饭和工作。在这里,我们带了一群“夜猫子”,试图在现实世界中使用简单、实用、非药物干预来重置他们习惯性的行为延迟时间。我们表明,大约两个小时的相位提前可以实现,同时伴随着抑郁和压力的主观评分的显著降低。此外,认知(反应时间)和身体(握力)表现的要素在“非最佳”时间显著改善,表现的日峰值出现在一天的早些时候。
相位提前
活动描记术分析显示,在实验组中,干预前后的活动描记术睡眠开始时间和醒来时间都有显著的进步。干预前后的睡眠持续时间、潜伏期和效率都保持相似,证实睡眠开始时间越早,睡眠潜伏期越长,睡眠持续时间越短。因此,干预的行为影响可以具体归因于睡眠时间的改变,而不是睡眠稳态的改变。为了支持活动描记数据,我们还发现褪黑激素发作(DLMO)的显著相位提前了近2小时(00:02至22:04小时)。与此同时,皮质醇觉醒反应的峰值时间也有类似的提前,从11:19移动到09:06。相角(测量DLMO和睡眠开始之间的时间)在干预前后也是一致的。通过使用金标准的昼夜节律相位标记,除了客观的活动图,这些结果表明在干预后在实验组中观察到了真正的昼夜节律相位提前。
由于光是昼夜节律系统的主要特征,因此它一直是治疗慢性睡眠障碍(如DSWPD)和情绪障碍(如季节性情感障碍)的主要选择之一。虽然在这项研究中没有特别进行控制光照,但参与者被要求更早醒来,并最大限度地暴露在晨光下,从而有助于一个阶段昼夜节律系统的进展。同时,观察到的较早的睡眠开始时间与减少例如来自房间照明和电子设备的夜间光暴露的指示相结合,可能有助于延迟DLMO和睡眠开始。食物摄入的时间也可能是影响相位提前的一个因素。有人认为进餐时间对昼夜节律系统,特别是与新陈代谢有关的外周时钟,有着影响。随着睡眠对食欲调节的重要性,研究发现,一顿富含碳水化合物的早餐可以提前CBT。自我报告的平均早餐时间(10:33到09:25小时)有了显著的提高,早餐的食用天数/周数也有所增加,尽管这并没有达到显著性。自我报告的午餐和晚餐的平均时间也是如此,这发生在干预后的更早时间,使我们能够确认遵守19:00后不吃晚餐的干预要求。在实验组而不是对照组中观察到的进餐时间的这些进步,可能有助于昼夜节律时间的进步,然而,由于食物的相移效应在这项研究中没有直接测量,它仍然是推测性的。
干预对心理健康和表现的影响
许多独立研究表明,睡眠延迟与精神健康下降(如抑郁)有关。针对睡眠和昼夜节律期也已成为神经心理障碍新疗法开发的焦点。干预后,我们发现实验组的抑郁和压力评分显著下降,表明心理健康状况有所改善。这与情绪障碍的类似趋势有关,POMS评分从10.33降至-2.89,尽管这没有达到统计学显著性。有趣的是,抑郁和压力因素显著降低,焦虑评分不受影响。虽然焦虑和抑郁是两种不同的疾病,有不同的诊断标准,但它们通常是并存的。然而,这些结果表明,每个因素都是独立影响的,表明与睡眠时间的可分离关系。这与文献表明的时间关系是一致的焦虑/抑郁和睡眠质量或数量的减少也是不同的(即焦虑通常在睡眠问题之前548,抑郁通常在睡眠问题之后。能够客观地分别探索这些因素并确定因果关系的方向将是这项工作未来的一个重要步骤,以确定该方法在改善心理健康方面的潜在临床用途。白天嗜睡,在这里用KSS测量,是与糟糕的表现和更高的出错风险相关的关键因素之一。嗜睡增加,导致注意力不集中,甚至微睡,被认为是每年记录的555起车辆相关事件的主要影响因素。无论是在临床环境中,还是在考虑现实世界中的表现/生产力时,能够减少日间嗜睡仍然是一个主要的556动机。这里我们表明,实验干预显著降低了08:00和14:00时的日间嗜睡。嗜睡在早上仍处于最高水平,尽管明显低于干预前。早晨这种接近两点的差异意味着从“有些困倦迹象”到“相当警觉”(KSS的分数是6到4)。KSS评分没有明显的日变化,类似于反应时间和握力的测量结果。此前,KSS评分已被证明与表现变量(如PVT)以及客观睡意有显著关联。因此,这种干预对那些通常更容易受嗜睡影响并需要高度警惕的职业,如空中交通管制、卡车驾驶和航空,可能是有用的,特别是因为事故风险已显示出日变化。
了解运动表现的日变化使得一些研究揭示了在一天中非最佳时间发生运动事故的高风险背后的原因,而另一些研究则考察了对运动员表现的影响。根据这些建议,我们现在展示了操纵夜猫子(LCPs)的这些日变化的潜力,产生一个相位提前,以创建一个在一天中较早发生的峰值性能的轮廓。实验组在“非最佳”早晨时间的反应时间(用PVT测量)和等长握力(用MVC测试测量)有显著改善,而对照组没有。
实验组还显示嗜睡和表现变量的日变化显著减少。这种幅度的降低与以前的研究一致,以前的研究表明夜猫子与云雀相比,表现差异的范围要大得多。反应时间和握力的日变化曲线反映了睡眠和昼夜节律时间的提前,握力峰值从20:00时移至15:21时,最快的反应时间出现在干预后12:30时,而不是20:00时。
07结论
在这里,我们展示了简单的非药物干预的能力,以提前夜猫子,减少精神健康和嗜睡的负面因素,以及操纵现实世界中的峰值表现时间。这些发现可以在许多不同的环境中产生相当大的益处。在普通人群中,其中很大一部分是夜猫子,这些发现可以提供一个简单的策略来改善精神健康和表现。在临床环境中,可以探索针对抑郁症和压力心理健康的进一步治疗,特别是针对昼夜节律紊乱,而不需要药物。这种干预也可应用于更为特殊的环境,如工业或体育部门,这些部门的重点是制定战略,以最大限度地提高生产率和优化绩效。尽管需要进一步的研究,但对于一个健康状况越来越差、精神健康状况越来越差、不断面临实现个人最佳表现的压力的社会来说,这仍然是一个令人兴奋的前景。
本文转载自其他网站,不代表健康界观点和立场。如有内容和图片的著作权异议,请及时联系我们(邮箱:[email protected])